来自中国科学院神经科学研究所、BGI Research、Monash University等团队,在 Science发表了题为
An opposing molecular gradient axis underlies primate cortical organization的研究。作者想回答的,是神经科学里一个非常经典、也一直没有真正统一的问题:灵长类大脑皮层,到底是沿着什么原则被组织起来的?
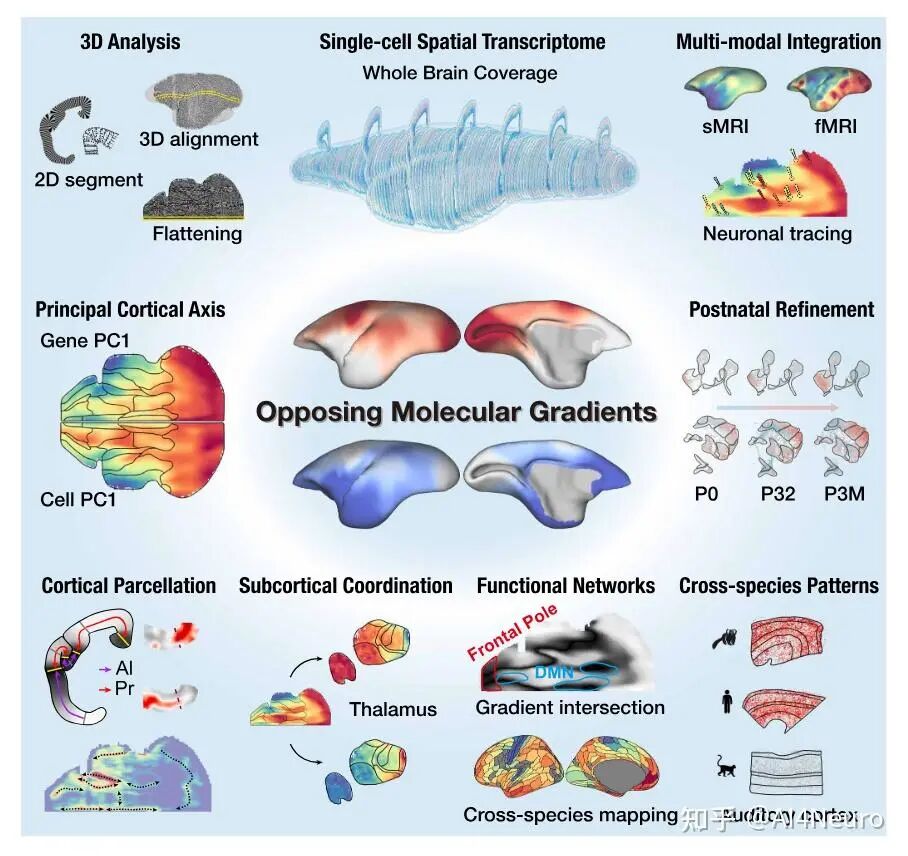
这个问题为什么重要?因为大脑皮层并不是一整块均匀的组织。它里面有初级视觉皮层、听觉皮层、躯体感觉皮层,也有和记忆、决策、社会认知相关的大量联络皮层。过去人们提出过不同解释:有的认为皮层是从更古老的皮层区逐渐展开、分化出来的;也有的认为初级感觉区才是更早、更稳定的“锚点”,其他皮层是在这些锚点周围继续扩展出来的。两种说法各有依据,但一直没有一个足够整体的框架把它们真正统一起来。 这篇研究的核心结论非常漂亮:这两种看起来互相竞争的理论,其实都只说对了一半。作者发现,灵长类皮层内部存在两股彼此相反的分子梯度:一股从古皮层/旁古皮层方向发出,另一股从初级感觉皮层方向发出,而广义的联络皮层正好位于这两股梯度的交汇地带。换句话说,大脑皮层并不是从单一中心向外展开,而更像是在两个端点的共同牵引下被组织起来的。更有意思的是,这个模式不只是一个抽象概念。作者把全脑空间转录组、单核测序、MRI、神经追踪放在一起后发现,这条“对抗分子梯度轴”不仅能解释皮层分区,还能解释丘脑-皮层对应关系、默认网络的位置、额极的特殊性,甚至还能解释为什么狨猴的听觉皮层在某些分子特征上比猕猴更像人类。
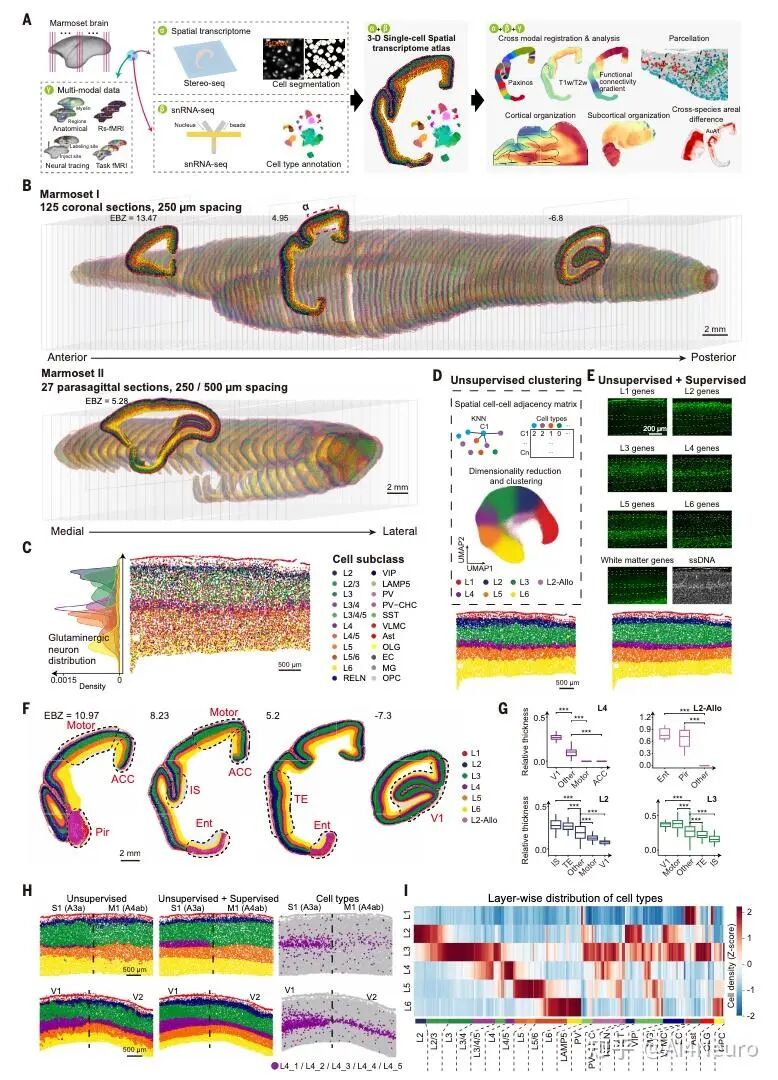
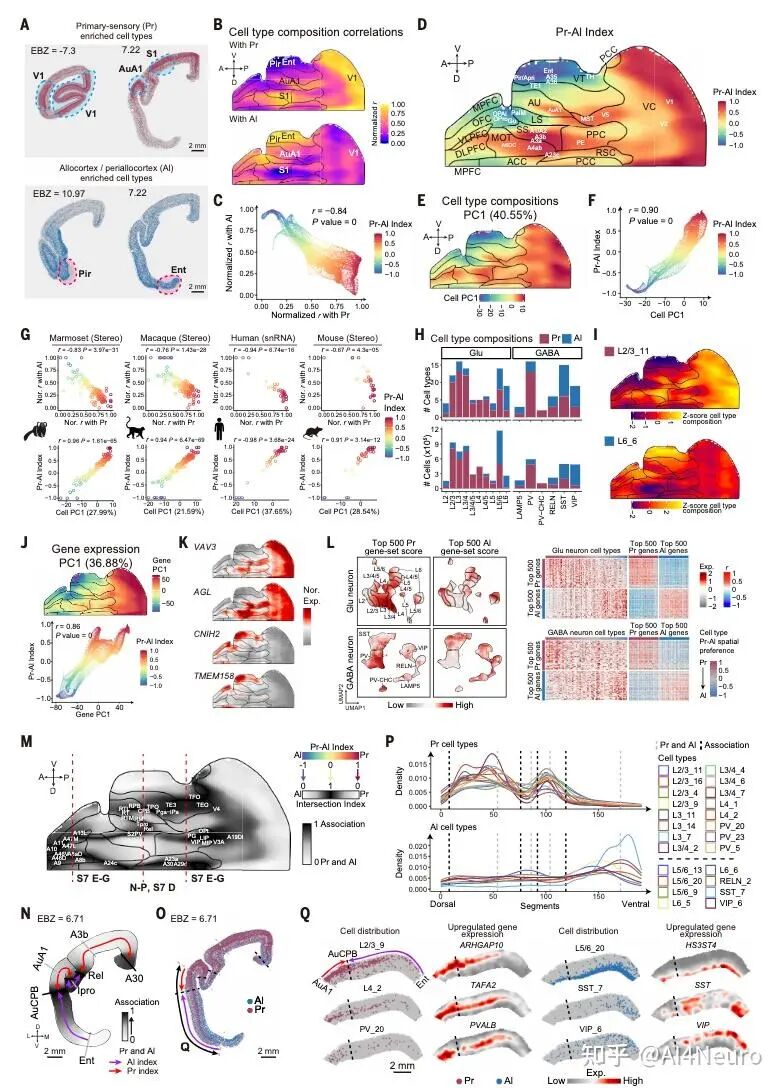
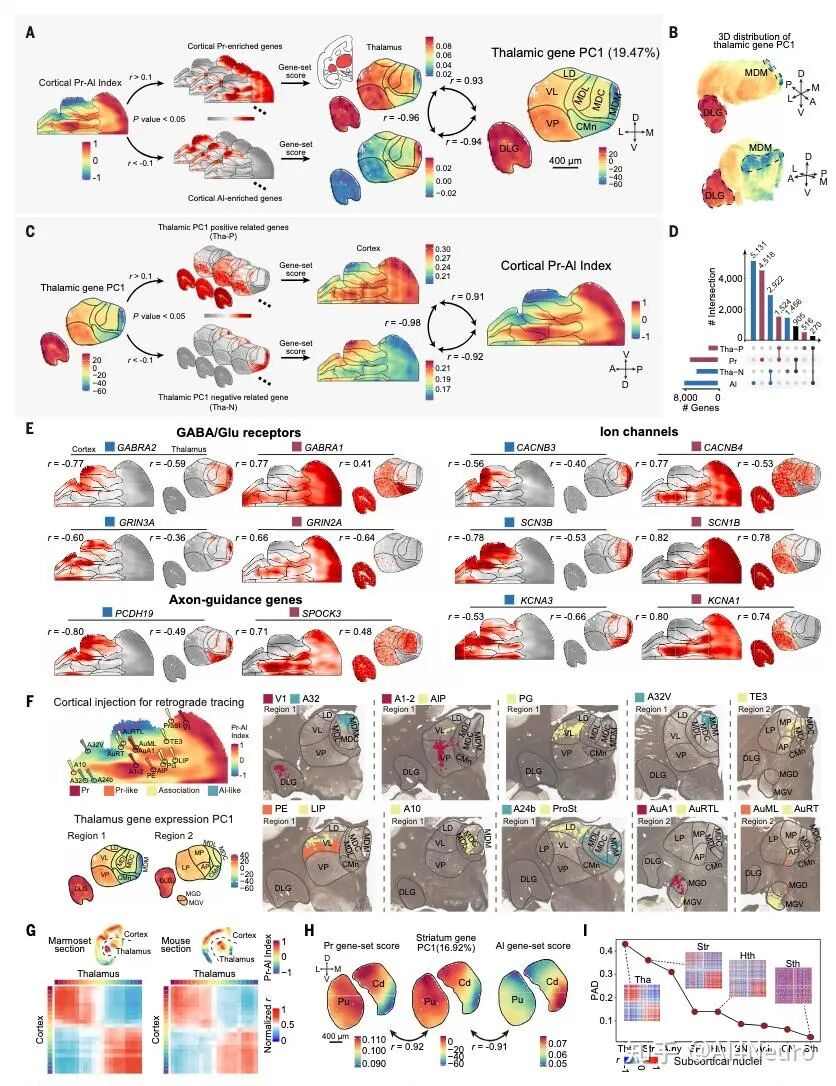
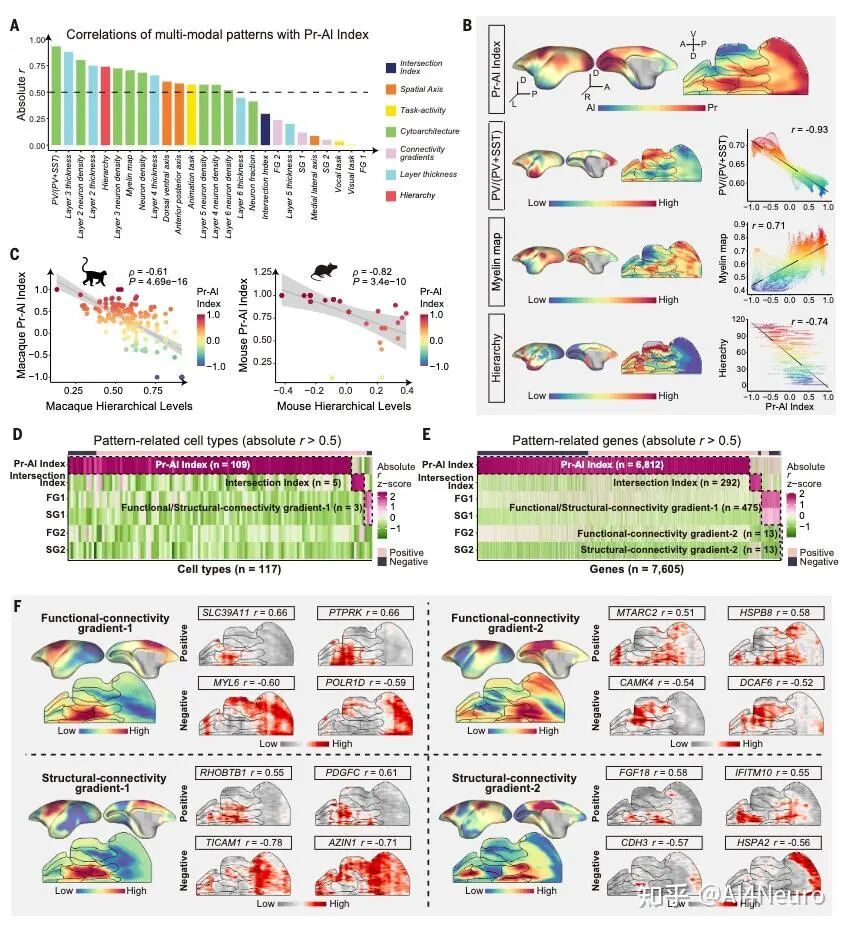
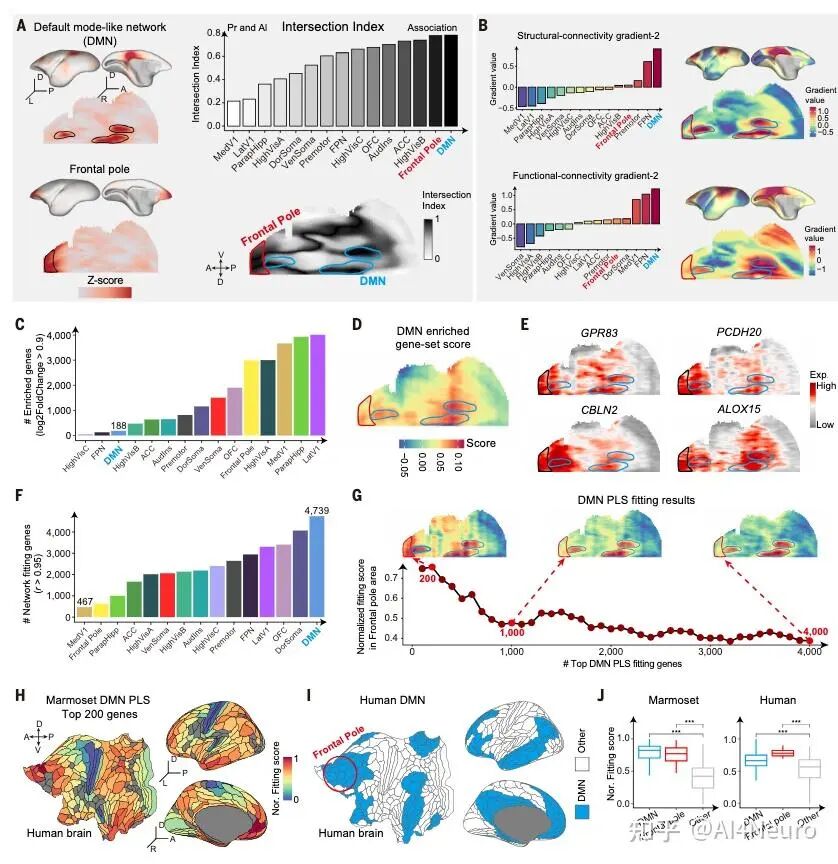
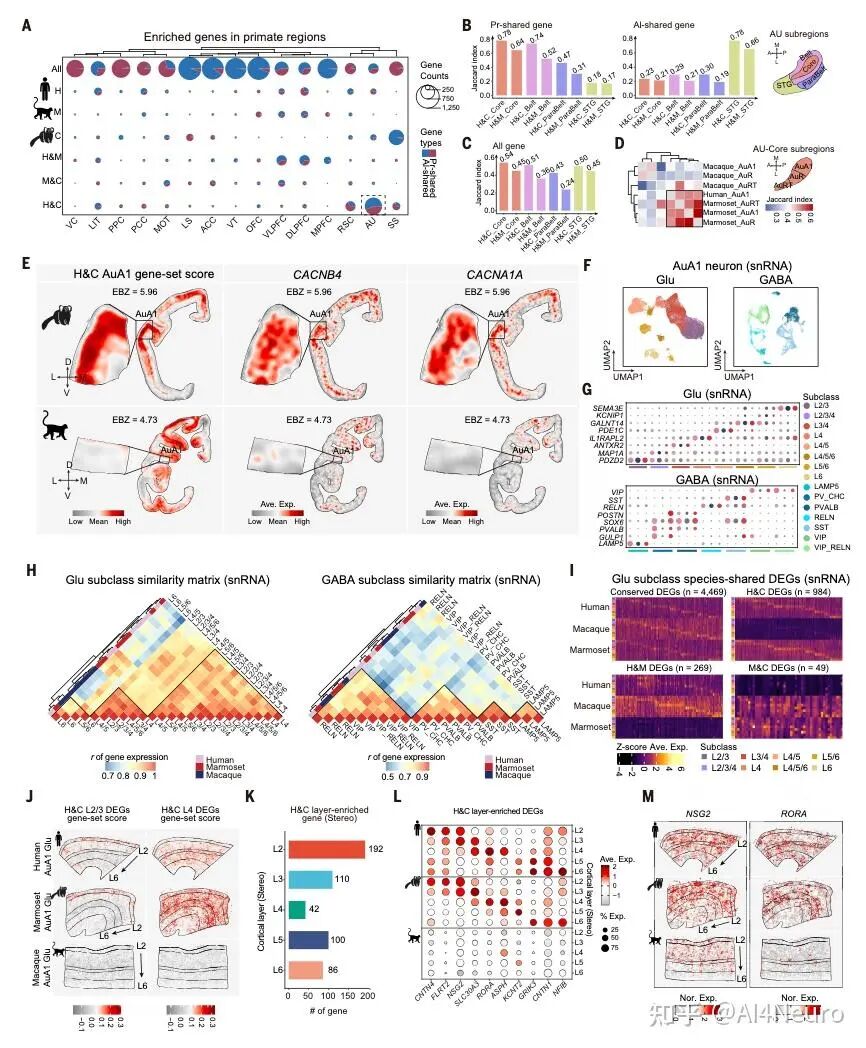
这篇论文最重要的一步,是先把争论双方都认真放进了数据里。作者选用的是狨猴。原因很现实:狨猴是灵长类,保留了很多关键的灵长类皮层特征,但它的大脑表面相对光滑,没有像人类和猕猴那样复杂的脑回折叠,因此更适合做整脑、精细、跨模态的空间整合。研究团队对狨猴做了全脑单细胞分辨率空间转录组,并结合单核 RNA 测序、MRI 和逆行神经示踪,最后建立了一套三维整合图谱。 他们先看不同细胞类型在皮层中的分布。结果发现,有一批细胞类型明显更偏向初级感觉皮层,比如视觉、听觉、躯体感觉相关区域;还有一批则更偏向古皮层/旁古皮层,比如梨状皮层、内嗅皮层这些更古老、和记忆及嗅觉系统关系更深的区域。 接着,作者把这些区域各自当作“参考端点”,对整张皮层图做相关分析。结果非常清楚:如果一个区域越像初级感觉皮层,它就越不像古皮层/旁古皮层;反过来也一样。两者呈现出很强的负相关。于是他们进一步构建了一个连续指标,把整张皮层放到“初级感觉端”和“古皮层端”之间。最后看到的并不是杂乱的拼图,而是一条非常稳定的组织轴线。 这一步真正厉害的地方在于,它没有简单说“哪种理论对”,而是告诉我们:两个端点都是真实存在的,而且它们共同定义了皮层的大尺度组织。 图1|灵长类皮层可以被放进一张从两端展开的分子地图里 研究者整合狨猴全脑空间转录组、单核测序、MRI 和神经追踪数据,构建了一个三维多模态图谱,并进一步在细胞类型和基因表达层面识别出皮层内部的大尺度组织轴,为后续分析皮层层次、分区和连接关系提供了统一坐标。 一旦把这条轴线建立出来,皮层里很多原来不好解释的区域关系就开始变清楚了。 最容易理解的是,两端当然分别对应两类最鲜明的皮层:一端更接近初级视觉、听觉、躯体感觉皮层,另一端更接近梨状皮层、内嗅皮层这类古老区域。真正有意思的是中间部分。作者发现,所谓“联络皮层”并不是简单介于两者之间的平均态,而是更像一个交汇区。它一方面不完全像初级感觉区,另一方面也不完全像古皮层,而是在两股梯度的碰撞和混合中形成了自己的位置。 从细胞组成上看,这个组织轴对应的是皮层细胞类型分布的主变化方向;从基因表达上看,第一主成分也高度贴合这条轴;更关键的是,这个模式并不只在狨猴里成立,作者在人类、猕猴、小鼠数据中都看到了相似关系。也就是说,这并不是某个物种偶然的结果,而更像是一条跨物种保存下来的基本组织原则。 这件事非常关键。因为它意味着:皮层的复杂功能分工,并不是先有很多离散区域,然后再勉强拼起来;相反,它们很可能首先共享一条连续的分子组织轴,然后才在这条轴上进一步细化出不同区域。 图2|初级感觉区和古皮层像两股相反的力量,而联络皮层正位于它们的交汇处 通过计算不同皮层区域与“初级感觉参考端”和“古皮层参考端”的相似性,研究者定义出一条连续的分子梯度轴。该轴同时体现在细胞类型分布和基因表达中,且在人类、猕猴、狨猴和小鼠中都能看到相似模式。 如果这条梯度轴真的是皮层组织的基本原则,那么一个很自然的问题就是:它到底是天生写好的,还是后天慢慢长出来的? 作者专门做了发育分析,取了狨猴在出生当天、出生后 32 天、出生后 3 个月和成年期的空间转录组与单核测序数据。结果发现,这条“初级感觉—古皮层”的对抗梯度在出生时就已经能看见雏形,但远远没有成年时那么清晰。出生后,相关基因集之间的分离程度会不断增强,梯度越来越锐利。 而且,这种增强主要发生在神经元里,尤其是谷氨酸能和 GABA 能神经元,而不是普遍出现在所有非神经元细胞中。这说明,皮层组织轴的成熟,很可能和出生后的感觉输入、神经回路活动以及区域间连接的逐渐建立密切相关。 这一步让整篇文章一下子有了更强的发育意味。它不是在说“大脑皮层有一条静态地图”,而是在说:这张地图从出生开始就存在,但它会在经验和回路成熟的推动下被不断打磨。 图3|皮层的对抗梯度出生时就有雏形,但真正清晰起来是在出生之后 研究者比较了不同发育阶段的狨猴数据,发现初级感觉端和古皮层端的分子差异会随着发育逐渐拉开,说明这条组织轴并非完全先天定型,而是在出生后持续被塑造和强化。 这篇研究还有一个特别实用的价值:它不仅提出了一条组织原则,还拿它去重新理解皮层分区边界。 传统脑图谱很多是基于细胞结构、髓鞘分布或者连接模式划分边界。但作者这里发现,只要去看细胞类型密度和基因表达沿着皮层位置的变化速度,就能找到一批变化特别剧烈的地方,而这些地方往往正好对应皮层区域之间的分界。 有些边界的特点是“很多细胞类型一起突然变化”,说明这里是一次比较集体的转折;但也有些边界总体看起来过渡平缓,却有少数关键细胞类型或者关键基因突然跳变。这意味着,皮层边界并不只有一种样子。有些是大张旗鼓地换区,有些则是由少数分子特征悄悄地划出线来。 更有意思的是,作者发现这些高变化区域并不全都能被现有图谱解释。也就是说,在传统分区方法之外,空间转录组和细胞图谱还可能揭示过去没被明确识别的新亚区。论文里就举了两个例子:一个在内嗅皮层内部,一个在初级运动皮层内部,都出现了清晰的分子分界信号。 这提示我们,大脑皮层的“地图”也许还没有画完。未来一些看起来已经很熟悉的区域,可能还会继续被拆出更细的新分区。 图4|皮层边界往往出现在梯度骤变的地方,而且有些新边界以前并没有被清楚画出来 通过追踪细胞类型密度和基因表达的变化速率,研究者识别出一系列变化最剧烈的区域,这些位置往往和已知皮层边界吻合,同时也提示了一些过去未被明确标出的潜在新分区。 如果这条分子梯度轴只是皮层内部的现象,那当然已经很有意思;但这篇文章更进一步的地方,是作者发现这套逻辑还延伸到了丘脑。 他们把皮层中与两端相关的基因集拿去看丘脑表达,结果发现丘脑内部也呈现出很强的对应分化:一部分区域更接近“感觉端”基因模式,一部分区域更接近“古皮层端”模式。再结合逆行神经示踪结果来看,初级感觉皮层更多连接到丘脑中一类区域,而更接近古皮层的皮层区则更多连接到另一类区域,中间的联络区对应的也是中间型丘脑区。 这意味着,皮层和丘脑之间并不是简单的“一个发信号、一个收信号”,而更像是共享了一条共同的分子组织原则。作者还把这个结果拿去和小鼠比较,发现狨猴皮层—丘脑之间的这种分子耦合比小鼠更强,提示灵长类在丘脑-皮层协同组织上可能走得更远。这个结果很漂亮,因为它让我们看到,大脑里大尺度组织原则不是只停留在皮层表面,而是贯穿到与皮层紧密互联的皮层下结构中。 图5|皮层和丘脑像是在沿着同一条分子坐标轴彼此对接 研究发现,丘脑内部的基因表达主轴与皮层的对抗梯度轴高度对应,而逆行示踪结果进一步说明,这种分子对应关系与丘脑—皮层连接拓扑是一致的,并且在灵长类中表现得比小鼠更明显。 为了判断这条“对抗梯度轴”到底是不是一条真正有普适解释力的组织原则,作者把它和一系列已知的大脑空间模式做了对照。 结果发现,它和PV/SST 中间神经元比例、髓鞘化程度、皮层层级结构都有很强相关性。特别是与层级结构的关系非常明显:越靠近某一端,越接近感觉加工早期区域;越靠近交汇区,则越接近更高阶、更综合的区域。 相反,它和某些结构/功能连接梯度之间的相关性反而没那么强。作者的解释也很有启发性:连接梯度是对全局连接模式做降维后的结果,虽然有用,但可能会把某些具体网络的分子特征“平均掉”。而这条分子轴则更直接地反映了皮层细胞和基因层面的组织。 这点很重要,因为它说明:大脑的功能网络当然重要,但如果想追问“这些网络为什么长成这样”,分子和细胞层面的连续组织轴可能更接近底层答案。 图6|这条分子轴不是孤立存在的,它和皮层层级、髓鞘化与中间神经元构成一起指向同一套组织框架 研究将对抗梯度轴与多种空间模式对照后发现,它与皮层层级、髓鞘分布以及 PV/SST 细胞比例等指标高度相关,说明它不是局部现象,而是能贯穿多种结构与功能表征的底层组织原则。 这篇论文还有一个很容易引发兴趣的点,就是它对默认网络(DMN)和额极(frontal pole)关系的处理。在人类大脑里,额极通常被看作默认网络的重要枢纽之一。但在狨猴中,额极和默认网络之间的功能连接并没有人类那么强。按常规思路,似乎很难把它们放在一起。但作者发现,如果不先看连接,而先看分子位置,它们都位于那条对抗梯度轴的交汇高点附近。 进一步分析后,作者发现狨猴的额极和默认网络在基因表达上高度相似。也就是说,尽管它们在功能连接上还没有完全整合到一起,但在分子身份上,已经带有相似特征。作者据此提出一个很有启发的想法:也许额极作为默认网络一部分的“分子身份”比它在连接上的整合出现得更早。 到了人类,这种本来就相似的分子底子,进一步发展成了更强、更完整的网络连接。 这其实是非常有想象力的一步。它让我们看到,脑网络的演化不一定先从连接开始,也可能先从分子身份和区域潜能开始。 图7|额极和默认网络也许先是在分子层面相似,后来才在演化中被更强地连成一个整体 在狨猴中,默认网络位于对抗梯度交汇最明显的位置,而额极虽然在功能连接上不如人类那样显著,却已经与默认网络共享高度相似的分子特征。这提示网络整合可能建立在更早出现的分子身份之上。 这篇文章最后一个很出彩的结果,是跨物种比较中的“反直觉发现”。按系统发育距离,猕猴当然比狨猴更接近人类。可作者在分析和这条梯度轴相关的一组共享基因时发现,在听觉皮层上,狨猴和人类的相似性反而高于猕猴。尤其是在听觉核心区,这种相似最明显。 进一步拆到细胞层面后,作者发现这种相似性主要体现在谷氨酸能神经元上。很多在人类和狨猴听觉皮层中共同富集的差异基因,与离子通道、轴突发生、突触成熟和神经回路可塑性有关;而这些基因在猕猴里就没有那么一致的模式。 作者给出的解释也很有意思:尽管狨猴和人类在演化上相隔很远,但两者都发展出了相对复杂的社会发声和声音交流行为,因此听觉皮层可能在某些分子机制上出现了趋同。也正因为这样,狨猴或许会成为研究复杂声音交流乃至语言起源神经基础的一个特别值得重视的模型。 图8|在听觉皮层上,狨猴和人类出现了出人意料的分子相似性 跨物种分析显示,虽然猕猴与人类系统发育关系更近,但在听觉核心区,狨猴与人类共享了更多与神经信号传递、轴突发生和突触成熟相关的基因表达特征,提示复杂声音交流可能推动了趋同的分子演化。 这篇 Science 最值得记住的,不只是“发现了一条梯度”,而是它改变了我们理解皮层组织的方式。过去我们很容易把皮层看成很多离散区域拼出来的地图:视觉区一块、听觉区一块、前额叶一块、默认网络一块。这样的分区当然有用,但它更像结果。今天这篇文章往前推进了一步,它告诉我们:在这些区域真正成为各自区域之前,皮层内部可能早就存在一条连续的、由两端共同拉开的分子组织轴。 正是这条轴,给后续的分区、连接、层级和网络留下了基本框架。 这也解释了为什么过去关于皮层起源的不同理论会长期并存。因为它们都抓住了一部分真实:古皮层端是真的,初级感觉端也是真的。问题不在于谁完全对、谁完全错,而在于它们其实共同构成了一条统一轴线的两端。 当然,这篇研究也有边界。作者自己提到,当前样本量还不足以系统讨论个体差异和性别差异;发育分析还主要集中在出生后阶段,真正想看这条梯度怎么在胚胎期出现,还需要更早的发育数据;而且,虽然文章为皮层新边界提供了强有力线索,但要真正建立新的标准分区图谱,还需要进一步的功能验证。但即便如此,这篇工作已经足够重要。因为它提供的不只是一个漂亮结论,而是一条能把皮层分区、丘脑对应、默认网络、额极演化、听觉跨物种特化都串起来的主线。 灵长类大脑皮层之所以能变得这么复杂,也许正是因为它从来不是从一个地方单独长出来的,而是在两股相反分子梯度的共同牵引下,被一步步组织成今天这个样子。 文献来源:Huang Z, Yang Q, Li S, et al. An opposing molecular gradient axis underlies primate cortical organization. Science 392, eaea2673 (2026). 文献连接:https://www.science.org/doi/10.1126/science.aea2673